(English version at the end of the article)
Naše znalosti o dinosaurech se za poslední léta v mnohém změnily. Ukázalo se, že to, co jsme si o jejich životě mysleli před 20 lety, bylo téměř všechno špatně. S vědeckým pokrokem se stále více vynořuje na povrch nová a až neuvěřitelně překvapivá tvář vládců druhohor. Evoluce dinosaurů se ubírala podivuhodnými směry od prapodivných morfologických struktur až po složité sociální chování a ve vědeckém světě zcela právem probíhá skutečná revoluce v pohledu na tyto tvory.

Morfologické
změny během ontogeneze klíčem k pestrému sociálnímu životu
Diverzita dinosaurů sahá od drobných forem velikých jako kočka až po opravdové giganty s mnohatunovou hmotností. V minulosti byla pozornost paleontologů věnována především pokusům o nalezení obrovských kosterních pozůstatků a muzea se předháněla v tom, které z nich vlastní větší exemplář. Když ale došlo na otázku, kde jsou mladé exempláře dinosaurů, odpověď prostě zněla, že neexistují. Všeobecně se předpokládalo, že mláďata vypadala jako zmenšené kopie svých rodičů, jak je tomu u dnešních plazů. Díky práci jednoho z nejúspěšnějších paleontologů posledních desetiletí, Jacku Hornerovi, se ale tato představa začala bortit. Ukazuje se, že dinosauři byly ve způsobu růstu podobní spíše ptákům.
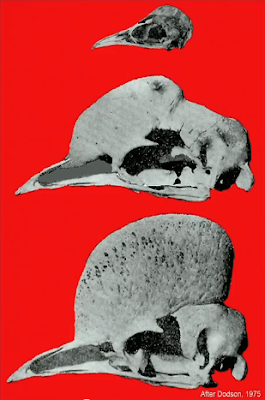
Novátorská myšlenka, že dinosauři možná rostli alometricky, Hornera napadla v jeho muzeu s názvem Museum of the Rockies během zkoumání lebek tří druhů pachycephalosaurů (Pachycephalosauria), dinosaurů s tlustou lebkou. Jednalo se o druhy Dracorex hogwartsia, Stygimoloch spinifer a Pachycephalosaurus wyomingensis. První z nich dosahuje nejmenšího vzrůstu a na zadní straně tlusté lebeční báze mu vyrůstají dlouhé kostěné výběžky. U druhého zvířete je velikost větší a útvary menší, zatímco třetí je naprosto postrádá a jeho lebeční báze je mnohem tlustší. Stavba lebky pachycephalosauridů s mohutnou frontoparietální zónou je mezi obratlovci naprosto unikátní. Bylo už publikováno mnoho prací zabývajících se funkcí této morfologické struktury, ale téměř žádná pozornost nebyla věnována jejím změnám během ontogeneze.


Horner si jednotlivé charakteristické morfologické struktury na lebkách všech tří druhů vyznačil jako landmarky a ukázalo se, že se ve skutečnosti jedná o tři ontogenetická stádia jednoho druhu. Dlouhé výběžky a absence zvětšené frontoparietální zóny charakterizují nejmladší jedince popsané původně jako druh Dracorex hogwartsia. Postupně s růstem dinosaura dochází ke vtlačování výběžků do lebeční báze až do stádia kostěných pupenů. Zároveň se ztlušťuje svrchní část lebky. Hypotéza byla podpořena také histologickou analýzou kostí vykonanou po jejich příčném rozřezání. Lebeční kosti mladého jedince mají pórovitou strukturu se silnou vaskularizací pro účel rychlého růstu, naopak u dospělců nalezneme velice denzní strukturu s Haversovými kanálky bez silné vaskularizace. Všechno zapadalo do skládanky – Dracorex je mládě, Stygimoloch subadult a Pachycephalosaurus dospělec. U druhého stádia byl dokonce objeven také intrafrontální šev, který zaniká až po srůstu frontálií v období, kdy jedinec dosáhne dospělosti. Ze tří druhů pachycephalosaurů se najednou stal jeden.

To ale nebylo zdaleka všechno. Dlouholeté zkušenosti s hledáním fosílií ve slavné americké geologické formaci Hell Creek přivedly Jacka Hornera na stopu dalšího omylu v paleontologii. Zaměřil se na hledání mláďat jednoho z nejlépe známých dinosaurů –Triceratops horridus. Patří k ceratopsidům, rohatým dinosaurům, kteří v období Křídy ve velkých počtech obývali především severní polokouli. Triceratops je znám více než sto let a za tuto dobu byly popsány jen 4 lebky nepatřící adultům. Jak se ale ukázalo, důvodem byla jenom skutečnost, že hledání mláďat nebyla věnována dostatečná pozornost. Horner díky své práci během krátkého časového období objevil několik lebek mláďat různého věku, které pak mohly být vzájemně srovnány. Poprvé v historii se dalo zkoumat, jak se měnila morfologie Triceratopse během jeho života.

Do výsledné analýzy vstoupilo 10 lebek od délky 38 cm až po 2 metry a byla nalezena až šokující alometrie v rámci 4 růstových stádií. Hlava mláděte připomíná svým celkovým vzezřením proporce lebek juvenilních jedinců dnešních zvířat (veliké oči, krátká lebka, redukované znaky adultů). Límec tvořený kostmi os parietale a os squamosum je plochý a s věkem se postupně prodlužuje a rozšiřuje. U mláďat se na jeho obvodu nacházejí epoccipitalia, kostěné výběžky, které se s přibývajícím věkem zanořují zpátky podobně jako výběžky mladých jedinců u Pachycephalosaurů. Také postorbitální rohy procházejí mohutnou proměnou. Nejmladší jedinci mají jen malé základy rohů, které se u subadultů prodlužují a posteriorně ohýbají. Až v dospělosti dojde k jejich narovnání a konečnému zahnutí anteriorně. U mláďat odděluje kost os supraoccipitale od sebe dvě kosti os exooccipitale a tvoří střechu mozkovny a otvoru foramen magnum. U subadultů se přesouvá postupně dál a u dospělců už je kompletně oddělena od foramen magnum a překrývá exooccipitália. Dalším důležitým znakem dospělých jedinců je podle Hornera vytvoření parietálních oken v kostěném límci. U Triceratopse nebyly zatím tyto okna prokázány. Návrh na zrušení platnosti dalšího druhu dinosaura na sebe nenechal dlouho čekat. Tentokráte se jedná o druh Torosaurus latus, od kterého byly doposud známé jenom pozůstatky dospělců s otvory na kostech tvořících límec. Po důkladném zkoumání lebek vyšlo najevo, že čím starší jedinec byl, tím měl tenčí os parietale v místech, kde se u Torosaura nacházejí již zmíněné otvory. Horner teď mohl prokázat, že Torosaurus ve skutečnosti představuje dospělého Triceratopse s plně vyvinutou hustou kostí bez silné vaskularizace a s Haversovými kanálky. Po odebrání vzorků z postorbitálních rohů byla u nově popsaných stádií opět nalezena struktura typická pro juvenily, naopak stádium Torosaura mělo jednoznačně strukturu typickou pro dospělce. Jeho práce tedy vedla k návrhu nechat „vyhynout“ už tři druhy dinosaurů.

Na základě nových poznatků můžeme očekávat revoluční změny našeho pohledu na ontogenezi dinosaurů i u dalších druhů. Předpokládá se dokonce, že možná až jedna třetina všech popsaných druhů ve skutečnosti představuje určitá ontogenetická stádia druhů jiných. Fascinujícím příkladem je případ Tyrannosaura rexe, nejobávanějšího a nejznámějšího dinosaura. Podobně jako u Triceratopse byly i u tohoto druhu po dlouhou dobu vzácností fosilní zbytky mláďat. V Cleveland Museum of Natural History je ale uložena lebka, která budí mnohé pochybnosti a byla základem pro Hornerovu hypotézu o zrušení validity dalšího druhu dinosaura. Oficiálně patří druhu Nanotyrannus lancensis, což je drobný a štíhlý bratranec obávaného Tyrannosaura. V důsledku předchozích objevů u pachycephalosaurů a Triceratopse se ale na scéně objevila otázka, jestli se ve skutečnosti nejedná o mladého jedince mnohem většího predátora. V jedné ze svých předchozích studií Horner pomocí osvědčené techniky zkoumání histologie kostí zjistil, že Tyrannosaurus rex rostl přibližně stejným tempem jako dnešní slon africký (Tyrannosaurus dosáhl dospělosti ve věku 20 let a slon ve věku 25 let). Znamená to tedy, že mladá zvířata pravděpodobně vedla velice aktivní život a morfologicky se v mnohém od dospělců lišila.

Názory na lebku uloženou v Clevelandském muzeu se doposud rozcházejí a je považována buď za mládě Tyrannosaura rexe, nebo za Nanotyranna. Jedním z hlavních argumentů proti první hypotéze je skutečnost, že zatímco menší z tyrannosaurů má v čelisti 17 zubů, ten nejmohutnější jenom 12 až 13. Je skutečně možné, že dinosauři během dospívání přicházeli o zuby? Jacku Hornerovi se povedlo něco podobného, jako u případu s rohatým Triceratopsem – našel během jedné sezony až 5 lebek obávaného predátora období Křídy, a navíc mezi nimi byla jedna výjimečná. Je větší než lebka Nanotyranna a menší než lebka dospělého Tyrannosaura rexe. V čelisti má 14 zubů, což je podle něj jasný důkaz, že tyrannosauři přicházeli s věkem o zuby a že zrušení platnosti dalšího druhu dinosaura je opodstatněné. Po srovnání lebek navíc vyšlo najevo, že u dospělců docházelo k vytvoření hrbolků na svrchních kostech horní čelisti a silnému zhrubnutí os postorbitale, což byly pravděpodobně jasné znaky sexuální zralosti. Jeden z největších predátorů v historii naší planety tedy započal svůj růst jako drobný subtilní lovec malých živočichů, který se od dospělců morfologicky velmi lišil.

Ne všichni odborníci se shodují v názorech na nové vnímání ontogeneze dinosaurů a případné rušení dříve popsaných druhů. Navržené zrušení validity Torosaura například vyvolalo zapálené reakce dalších paleontologů a vlnu nespokojenosti i v řadách laiků, navíc situace není doposud uzavřena. Důkazů pro existenci bizarních transformací ale pořád přibývá. Jednou z dalších skupin dinosaurů s ohromnou diverzitou různých kostěných útvarů, které vznikly patrně jako výsledek sexuální selekce, jsou hadrosauři. V budoucnu dojde nejspíš právě u těchto sociálních býložravců ke zrušení platnosti několika druhů. Zatím se pozornost věnuje především kraniální anatomii, ale studium postkraniálních částí kostry může odhalit další překvapivé informace o tom, jak dinosauři rostli. Je velice pravděpodobné, že většina dinosaurů během svého růstu procházela několika ontogenetickými stádii.

K jakému účelu ale sloužila existence z energetického hlediska nákladných morfologických struktur? Odpověď je prostá – k sociálnímu životu. Pro studium evoluce chování dinosaurů nám nejlépe poslouží ptáci, jejich přímí potomci. Jak bylo experimentálně zjištěno, nalezneme u nich pořád geny zodpovědné za tvorbu zubů nebo ocasu, jejichž exprese je pouze potlačena v důsledku evolučních adaptací. Podobně jako dinosauři, i mnozí ptáci disponují morfologickými strukturami, které jim pomáhají komunikovat mezi sebou v rámci societ. Tyto struktury se navíc s věkem vyvíjejí. Příkladem můžou být kasuáři nebo zoborožci, u kterých si mláďata až do poměrně pozdního věku uchovávají juvenilní znaky. Říkají: „Jsem mládě, ne hrozba.“ Naopak jedinec nesoucí znaky dospělce může představovat soka, nebo naopak sexuálního partnera. Různé morfologické struktury byly pravděpodobně podobně jako u ptáků pestře zbarveny a doplňovaly složitou mozaiku jejich sociálního chování.

Our knowledge of dinosaurs have differed a lot in the last few decades. It became clear, that almost everything we thought about them 20 years ago is wrong. The new and surprising face of kings of Mesozoic period is coming up. The evolution of dinosaurs went through the extraordinary paths from stunning morphological structures to the complex social behaviour. We live in the age of new dinosaur revolution.

The morphological changes during ontogeny as the key to the variable social life
The dinosaur diversity reaches a huge scale from the tiny ones in the size of a cat to the true giants with weight of tons. In the past the strongest attention was focused on searching for the biggest fossils and museum were competitors in possesing as huge bones as possible. But when it came to the question of juveniles, almost everybody assumed that they don't exist. The overall theory was, that youngsters looked as the smaller copies of their parents, same as in recent reptiles. Thanks to the work of one of the most successful paleontologists in last decades, Jack Horner, this opinion about dinosaur ontogeny started to fall apart. It appears, that dinosaur grew up a lot more similarly to birds.
The new idea, that dinosaurs grew with allometric speed, came to Horner in the Museum of Rockies during examination of skull of three species of pachycephalosaurs (Pachycephalosauria). These were Dracorex hogwartsia, Stygimoloch spinifer and Pachycephalosaurus wyomingensis. The first one is the smallest and on the posterior part of thick skull base it has a prominent bone thorns. The second species is bigger and its thorns are smaller, the third one completely lacks them and the skul base is much stronger. The skull morphology of pachycephalosaurs with the robust frontoparietal zone is unique in vertebrates. Many studies were published on the function of these structures, but almost no attention was focused on their ontogeny.


Horner highlighted the characteristic structures on skull as landmarks and it showed out, that they represent a three ontogenetic stages of one species. The long horns and the absence of enlarged frontoparietal zone are characteristic for the youngest individuals described originally as Dracorex hogwartsia. During growth the horn are getting back into the main bone and they end as pyramidal nodes. The upper part of skull base is getting thicker. The hypothesis was also supported by the histological analysis of bones done after their transversal cutting. The skull bones of young individual have a porous structure with the strong vaskularization for the quick growth, in adults we find a very dense structure with the Haversian tissue and without stronger vaskularization. Everything matched into the puzzle - Dracorex is a youngster, Stygimoloch is a subadult and Pachycephalosaurus is and adult animal. In the second one, even the intrafrontal suture was found, which dissapears after the connection of frontalia and indicates a stage of growth in young individuals. From three species of pachycephalosaurs there was suddenly only one.


But this was not the end. The long lasting experiences with the hunting of fossils led Jack Horner to the next mistake of paleontology. He focused on the one of the best known dinosaurs -Triceratops horridus. It belongs to ceratopsians, the horny dinosaurs mainly from the northern hemisphere of Cretaceous. Triceratops is known more than 100 years and only 4 skull of young animals have been described ever since. However, as it became clear, this was caused only by the small attention focused on searching for small skulls. Horner found a few skull in the short period of time and they could be compared together. For the first time in the history the ontogeny of Triceratops could be examined.

10 skull reaching from 38 cm to 2 meters were included in the analysis and there was a shocking allometry found in the 4 growth stages. The head of young animal resembles the proportions of recent juveniles in animal kingdom (big eyes, short skull, reduced adult features). Frill build from os parietale and os squamosum is flat and during ontogeny it becomes longer and wider. There are epoccipitals (bony elements) at the edge of the frill in youngsters and they are gradually reduced and get smaller into the main bone as in young Pachycephalosaurs. Also the postorbital horns go through a big changes. The smaller ones have only the base of horns, which get longer and in subadults they curve posteriorly. After reaching adulthood they go straight again, but finally they curve anteriorly. In juveniles, os supraoccipitale separates two bones os exooccipitale from each other and forms a roof for the braincase and foramen magnum. In subadults it moves back and in adults it is completely separated from foramen magnum and is over exooccipitals. The next key feature of adults is according to Horner creating of parietal fenestra in the frill. In Triceratops, no parietal fenestra were found before. But the suggest for canceling another valid dinosaur species was just waiting. This case is about Torosaurus latus, which is known only from adult with such fenestra in their frill. After the complex examination of frill found by Horner it became clear, that the more old the animal the more thin the frill was in the place of os parietale. Horner could prove, that in fact Torosaurus is an adult Triceratops with the fully developed bone without strong vaskularization and Haversian tissue. It was proved after the examination of fragments of postorbital horns from recently discovered ontogenetic stages, where the bone structure matched for juveniles. In Torosaurus skulls the bone was typical for adult animal. Thus, the work of Jack Horner led to proposal to cancel 3 species of dinosaurs.

According to the new findings we can expect revolutionary changes in our point of view on dinosaur ontogeny. It is even assumed, that almost one third of dinosaur species in fact represent ontogenetic stages of one different ones. A fascinating example happened in the case of Tyrannosaurus rex, the most fearsome and best known dinosaur. Similarly to Triceratops, the fossils of juveniles were very rare in the past. But in Cleveland Museum of Natural History is one skull, which triggered doubts about existence of another dinosaur species and was a basestone for Horner and his hypothesis. Oficially it belongs to Nanotyrannus lancensis, a smaller and faster cousin of Tyrannosaurus. After the discoveries in pachycephalosaurs and Triceratops, question about juveniles of huge predator emmerged. In one of his previous studies Horner proved, that Tyrannosaurus rex grew in a similar speed to african elephant (Tyrannosaurus reached adulthood in 20 and elephant in 25 years). It basically means, that young animals probably led a very active life and their morphology was very different from adults.

Two hypotheses about classification of skull from Cleveland museum assign it to young Tyrannosaurus rex or Nanotyrannus. The main argument against the first one is the fact, that the smaller dinosaur has 17 teeth in the jaw, the bigger one only 13 or 12. Is it really possible, that dinosaur have lost their teeth during ontogeny? Jack Horner succeded in the same project as with Triceratops -he had found 5 skull in one season. And one of them was very unique. It is bigger than skull of Nanotyrannus and smaller than skull of adult Tyrannosaurus. It has 14 teeth in the jaw, which is a clear evidence for Horner, that tyrannosaurs have lost their teeth when growing and the suggestion of canceling a validity of another dinosaur species is correct. After the comparation of skull it appeared, that in adults there were protuberances on the upper bones of skull and a thickening of os postorbitale happened, which is probably a signal of sexual maturity. One of the biggest predators in the planet's history started its life as a small and subtle hunter of small animals, very different from adults.


But for what reason should exist the energetically costly morphological structures? The answer is simple -for the complex social life. To study dinosaur behaviour, we have to focus on their direct descendants, birds. There were exmeriments, which proved that genes responsible for the growth of teeth or tail vertebrae are still present in birds, their expression is just supressed because of other evolutionary adaptations. Similarly to dinosaurs, birds have many structures, which help them to communicate with each other. And finally, these are developing during ontogeny. The good example can be cassowaries aor hornbills. Their chicks retain the juvenile look for the long time and this basically sais „I am a juvenile, not a threat.“ On the other hand, adult features and structures are signals of enemy or sexual mate. These morphological structures were probably also very colourful in dinosaurs and were part of the living in societes.
Zdroje:

Zdroje:
HORNER J. R. & GOODWIN M. B. 2009: Extreme Cranial Ontogeny in the Upper Cretaceous Dinosaur Pachycephalosaurus. PLoS ONE 4(10): e7626. doi:10.1371/journal.pone.0007626.
HORNER J. R. & GOODWIN M. B. 2006: Major cranial changes during Triceratops ontogeny, Proc. R. Soc. B 7 (273): 2757-2761. doi: 10.1098/rspb.2006.3643.
SCANNELLA J. B. & Horner J. R. 2010: Torosaurus Marsh, 1891, is Triceratops Marsh, 1889 (Ceratopsidae: Chasmosaurinae): synonymy through ontogeny, Journal of Vertebrate Paleontology 30 (4): 1157–1168.
WITMER L. & M. RIDGELY R. C. 2010: The Cleveland tyrannosaur skull (Nanotyrannus or Tyrannosaurus): new findings based on CT scanning, with special reference to the braincase, Kirtlandia, 57:61–81.
HORNER J. R. & PADIAN K. 2004: Age and growth dynamics of Tyrannosaurus rex, Proc Biol Sci., 271(1551): 1875–1880.





















.jpg)












.jpg)






