
(English version at the end of article)
Čeleď Cheirogaleidae je mezi lemury zřejmě nejméně přehledná. Patří sem 5 rodů, z nichž dva jsou zatím docela velikým oříškem pro fylogenezi v rámci této skupiny. Tyto drobná, maximálně něco přes půl kila vážící zvířata se vyznačují nočním způsobem života, který z nich dělá opravdu těžké objekty na zkoumání. O mnohých druzích se doslova ví jenom to, že existují. V důsledku ohromné úrovně mikroendemismu jsou na určitém malém území objeveny, ale pro bližší výzkum stylu života nezbývá prostor (je na čase to změnit). Jako by toho nebylo málo, makiovití nezanechali po sobě žádný fosilní materiál, což se snad v budoucnu podaří změnit (vzhledem k jejich křehkým kostem je to ale dost nepravděpodobné). Studium evoluce tajuplných makiů je tedy velice neprobádanou oblastí a musíme se tedy držet jen toho, co zatím víme o žijících druzích.

Mezi makiovité patří nejmenší primát světa -Microcebus berthae. Obecně došlo u této čeledi k několika důležitým adaptacím, díky kterým mohli využít jiné ekologické niky než další skupiny lemurů. Především zaujme jejich malá velikost, noční způsob života a dlouhý ocas sloužící k manévrování. Oči jsou vybaveny přídavnou vrstvou buněk za sítnicí (tapetum lucidum), která umožňuje pohltit co nejvíc světla během tmavé noci. Mají základní znaky poloopic, jako zubní hřeben z řezáků nebo čistící dráp na druhém prstu zadní nohy, ale nalezneme u nich třeba nezvykle velké špičáky. Jsou to omnivoři schopní využít k obživě jak rostlinnou, tak živočišnou potravu. Poměr jednotlivých složek se ale druh od druhu mění a například druhy s velikým podílem mízy v potravě mají redukované mandibulární svalové adduktory a také oblasti, kam se upínají (koronoidní processus).
Poslední obsáhlá a komplexní studie ohledně molekulární stránky evoluce Cheirogaleidae je z roku 2001, což ukazuje, jak málo toho o trpasličích lemurech víme. Navíc v té době bylo popsáno jen 9 druhů, zatímco dnes jich známe 31! Přesto se zběžně na její výsledky podíváme. Už v té době byly odhaleny uvnitř čeledi 2 různé karyotypy. U rodu Phaner platí 2n =46, u ostatních čtyř 2n=66. Tenhle rod se zřejmě oddělil od společného předka této čeledi jako první, soudě podle odvozených morfologických znaků (Tattersall and Schwartz, 1974). Naopak imunologická data z albuminu a transferrinu spojují Phaner a Cheirogaleus dohromady a jako první diverzifikující rod se jeví Microcebus. Analýza dále odhalila překvapivou bazální divergenci rodu Cheirogaleus a sesterský vztah Microcebus a Mirza. Rod Allocebus s pouze jedním druhem byl tehdy velice málo prozkoumán a jeho pozice nebyla vůbec dořešena. Už v té době na základě mtDNA autoři předpokládali, že dojde k popisu nových druhů u rodu Microcebus. Dohromady nám vyjde, že Phaner a Cheirogaleus (možná společně s Allocebus) byli součástí adaptivní radiace a tvoří sesterskou skupinu k Microcebus a Mirza.
Cheirogaleidae se zřejmě odštěpili od hlavní fylogenetické linie lemurů jako druzí hned po Daubentonidae s jediným zástupcem, ksukolem ocasatým. Tito drobní primáti jsou často označováni za velice bazální skupinu, která si zachovala vzhled dávných předků všech primátů. Z evolučního hlediska možná vypadají primitivně, například jejich sociální vazby jsou ale velice rozmanité a najdeme například jak solitérní, tak velice společenské druhy. Typickým solitérem je Mirza coquereli, Phaner furcifer a Cheirogaleus medius žijí v jednotlivě rozptýlených párech se svými potomky v přesně vymezených územích, u Microcebus murinus tvoří samice skupiny ukládající se společně k spánku. Velikou roli při evoluci sociálních struktur sehrálo prostředí (dostupnost potravy, predátoři, kompetice mezi jedinci).

Kvůli nedostatečnému poznání fylogenetických vztahů u jednotlivých druhů čeledi se na každý z nich podíváme jednotlivě. Analýzy mitochondriální DNA naznačují existenci dvou skupin v rámci rodu Microcebus -východní a západní. Kvůli extrémní diverzifikaci se zde dají čekat objevy nových druhů na dosud neprobádaných lokalitách.
Rod Allocebus
Allocebus trichotis

Druh byl dlouhou dobu považován za vyhynulý a znovuobjevený až v roce 1989. Předpokládalo se, že je kriticky ohrožen, ale jeho výskyt byl už prokázán na více místech severovýchodního Madagaskaru (deštné lesy). Typickým znakem je dlouhá srst na uších. Druhé a třetí svrchní moláry jsou kaniniformní a první horní řezák je zvětšen. Tvar uší se podobá spíše kombám než lemurům z čeledi Cheirogaleidae. Jazyk je dost dlouhý ve srovnání s ostatními druhy čeledi. Je to typický noční druh, který přes den přespává v dutinách stromů. V období sucha zvyšuje svou tělesnou hmotnost o tukové zásoby. Rodiče ve svém teritoriu trpí i svá mláďata, jeho velikost je ale podle nových výzkumů mnohem větší než u jiných drobných lemurů. Zatím se nenašlo přesvědčivé vysvětlení, i když byla navrhována potrava sestávající převážně z hmyzu (někteří autoři překvapivě uvádějí jako hlavní zdroj potravy mízu stromů) nebo sezónní změny v chování. Ani při spánku v dutinách se Allocebus nechová nesnášenlivě. Evolučně je to určitě alespoň samostatný rod, i když se dalo uvažovat také o zařazení do samostatné čeledi (dnes ale díky molekulárním datům řadíme tenhle druh hluboko mezi Cheirogaleidae).

Rod Mirza
Mirza coquereli

Rod Mirza byl původně řazen do Microcebus. Svou velikostí a váhou až do 300 g a způsobem života se ale liší. V suchém období nehibernuje, ale zůstává aktivní a živí se především hmyzem řádu Homoptera. V jiných obdobích roku se krmí také ovocem nebo květy. Typicky solitérní druh, u kterého byl zjištěn pohlavní dimorfismus, což je u lemurů docela veliká vzácnost. Samci jsou větší a velikost jejich varlat se v průběhu roku mění (páří se s více samicemi). Druh se vyskytuje jenom v suchých lesích západní části ostrova (severní areál je dnes řazen k jinému druhu). Uši jsou veliké a málo ochlupené. Samice mají dva páry bradavek, jedny pektorální a jedny abdominální. Jedinci obou pohlaví si staví několik hnízd ve svém domovském okrsku, které slouží jako úkryt před predátory. Většinu času stráví tenhle druh osamoceně, ale malá část pozůstává ze sociálních interakcí jako vzájemné čištění nebo vokalizace.

Mirza zaza
Druh byl popsán teprve v roce 2005 na základě studia chování a morfologie. Genetické analýzy jeho validitu potvrdili, jižní druh byl od severního oddělen zhruba 2 miliony let. Mirza zaza je celkově menší, má menší ušní boltce, kratší ocas a špičáky. V mitochondriálním genomu (cyt b) se liší v 3,51 %. Severní druh je sice menší, ale samci mají mnohem větší varlata, což ukazuje na vyšší promiskuitu. Celkově je to zřejmě největší velikost varlat v poměru k tělu u primátů. Druh se liší i behaviorálně -v hnízdech na rozdíl od Mirza coquereli tráví hodně času i více jedinců pohromadě (byli zjištěni dokonce 4 dospělí samci v jednom hnízdě).

Rod Microcebus
Microcebus murinus

První popsaný (1777) a dlouho jediný známý druh rodu Microcebus. Trpasličí makiové patří mezi kryptické druhy žijící na malém území a tenhle druh je mezi nimi výjimkou -vyskytuje se na docela veliké ploše západního Madagaskaru v suchých lesích. Jsou to typičtí noční primáti s velikými očnicemi a dlouhým ocasem. Na tváři a v okolí genitálií najdeme žlázy sloužící k pachové komunikaci mezi jedinci. Samice jsou větší než samci a druh jako takový je největším v rámci rodu Cheirogaleus s váhou až 70g. V suchém období hibernuje (torpor) při rapidním snížení životních funkcí a teploty až na 7 °C, zatímco v ostatních částech roku se krmí rozličnou potravou od listů a květů až po různý hmyz. Samice jsou vůči samcům dominantní a mnohem snášenlivější k jedincům stejného pohlaví. V období estra není vagína samic pokryta srstí jako obvykle a u jinak samotářských samců dochází k vytvoření hierarchie. Dominantní samec ale nemá vůbec jisté všechna privilegia, protože samice se páří s více samci a dochází tedy k jakémusi „závodu ve zbrojení“ mezi pohlavími. Zatímco dominantní samci jsou většinou mohutní a snaží se vytvořit převahu nad ostatními, samice mají spíše tendenci vnášet do populace více genetické variability, tedy projevuje se „efekt vzácných samců“. Ke komunikaci využívají tito lemuři celou řadu akustických (některé nad hranicí lidské slyšitelnosti) i olfaktorických signálů. Až studium genetické variability u tohoto druhu odhalilo existenci mnoha dalších a z rodu s jedným druhem se náhle stal rod se třemi desítkami druhů.

Microcebus griseorufus

Tento druh se vyskytuje jižněji než předchozí a obývá tu nejsušší krajinu ostrova. Dorůstá menších rozměrů, má poněkud světlejší srst a mohutnější lebku v poměru k tělu. Studie v terénu navrhovali na základě morfologických odlišností vytvořit v rámci populace tohoto druhu druhy dva, analýza mitochondriální DNA ale ukázala, že se zde nenachází rozdíly mezi dvěma morfotypy. K takové nevídané plasticitě dochází v oblastech, kde makiové obývají více mikrohabitatů (v místech se společným výskytem s předchozím druhem se podobná variabilita neprokázala). Jako noční zvířata tito primáti nepoužívají vizuální komunikaci jako primární zdroj informací při sociálních interakciích, což může vysvětlovat existenci dvou morfotypů. V budoucnu bude ale potřeba připojit do analýzy také jaderné geny a možná nás čekají překvapivé výsledky. Zatím je ale Microcebus griseorufus jednotným druhem, o kterém se stále mnoho neví.

 |
| Dva morfotypy nalezené na lokalitě Beza Mahafaly patří k jednomu druhu |
Microcebus ravelobensis
 Pro změnu druh ze severní části Madagaskaru. Srst je rezavá a velice charakteristická. Vyskytuje se sympatricky s Microcebus murinus, rozlišíme jej ale podle tenšího ocasu, ve kterém neukládá nikdy tukové zásoby, podle delších zadních nohou a barvy. V důsledku délky zadních nohou se na rozdíl od Microcebus murinus pohybuje po větvích skákáním, ne lezením. Oba druhy se dále liší v načasování estra samic a Microcebus ravelobensis je agresivnější co se týče sociálních interakcí. Dále byla zjištěna potravní speciace a to hlavně u samic. U Microcebus ravelobensis je mnohem menší tendence zvyšovat podíl ovoce v jídelníčku a celkově dva druhy využívají stejné zdroje jen v období sucha. I tehdy ale nedochází ke kompetici kvůli jinému způsobu lokomoce (skákání a lezení).
Pro změnu druh ze severní části Madagaskaru. Srst je rezavá a velice charakteristická. Vyskytuje se sympatricky s Microcebus murinus, rozlišíme jej ale podle tenšího ocasu, ve kterém neukládá nikdy tukové zásoby, podle delších zadních nohou a barvy. V důsledku délky zadních nohou se na rozdíl od Microcebus murinus pohybuje po větvích skákáním, ne lezením. Oba druhy se dále liší v načasování estra samic a Microcebus ravelobensis je agresivnější co se týče sociálních interakcí. Dále byla zjištěna potravní speciace a to hlavně u samic. U Microcebus ravelobensis je mnohem menší tendence zvyšovat podíl ovoce v jídelníčku a celkově dva druhy využívají stejné zdroje jen v období sucha. I tehdy ale nedochází ke kompetici kvůli jinému způsobu lokomoce (skákání a lezení).

 Pro změnu druh ze severní části Madagaskaru. Srst je rezavá a velice charakteristická. Vyskytuje se sympatricky s Microcebus murinus, rozlišíme jej ale podle tenšího ocasu, ve kterém neukládá nikdy tukové zásoby, podle delších zadních nohou a barvy. V důsledku délky zadních nohou se na rozdíl od Microcebus murinus pohybuje po větvích skákáním, ne lezením. Oba druhy se dále liší v načasování estra samic a Microcebus ravelobensis je agresivnější co se týče sociálních interakcí. Dále byla zjištěna potravní speciace a to hlavně u samic. U Microcebus ravelobensis je mnohem menší tendence zvyšovat podíl ovoce v jídelníčku a celkově dva druhy využívají stejné zdroje jen v období sucha. I tehdy ale nedochází ke kompetici kvůli jinému způsobu lokomoce (skákání a lezení).
Pro změnu druh ze severní části Madagaskaru. Srst je rezavá a velice charakteristická. Vyskytuje se sympatricky s Microcebus murinus, rozlišíme jej ale podle tenšího ocasu, ve kterém neukládá nikdy tukové zásoby, podle delších zadních nohou a barvy. V důsledku délky zadních nohou se na rozdíl od Microcebus murinus pohybuje po větvích skákáním, ne lezením. Oba druhy se dále liší v načasování estra samic a Microcebus ravelobensis je agresivnější co se týče sociálních interakcí. Dále byla zjištěna potravní speciace a to hlavně u samic. U Microcebus ravelobensis je mnohem menší tendence zvyšovat podíl ovoce v jídelníčku a celkově dva druhy využívají stejné zdroje jen v období sucha. I tehdy ale nedochází ke kompetici kvůli jinému způsobu lokomoce (skákání a lezení).
Microcebus tavaratra
 Druh ze samého severu ostrova, kde obývá deštné lesy. Barva srsti pozůstává ze dvou nebo až tří různých, ale nejsignifikantnější je tmavý pruh táhnoucí se středem hřbetu. Na hlavě si lze všimnout tmavé vibrisy, které jsou u jiných druhů většinou bledé. Je jedním z druhů, o kterých se mnoho neví.
Druh ze samého severu ostrova, kde obývá deštné lesy. Barva srsti pozůstává ze dvou nebo až tří různých, ale nejsignifikantnější je tmavý pruh táhnoucí se středem hřbetu. Na hlavě si lze všimnout tmavé vibrisy, které jsou u jiných druhů většinou bledé. Je jedním z druhů, o kterých se mnoho neví.


Microcebus sambiranensis
 Druh byl objeven v roce 2000 na severu Madagaskaru při průzkumu v rezervaci Ankarana. Základní barva těla je skořicově hnědá. Srst na ocase je dlouhá a kolem očí vytváří tmavý prstenec. Stejně jako předešlý druh má tmavé vibrisy. Další z těch, kteří teprve čekají na důkladné prozkoumání. Díky výskytu v chráněné oblasti je výzkum poměrně slibný do budoucnosti.
Druh byl objeven v roce 2000 na severu Madagaskaru při průzkumu v rezervaci Ankarana. Základní barva těla je skořicově hnědá. Srst na ocase je dlouhá a kolem očí vytváří tmavý prstenec. Stejně jako předešlý druh má tmavé vibrisy. Další z těch, kteří teprve čekají na důkladné prozkoumání. Díky výskytu v chráněné oblasti je výzkum poměrně slibný do budoucnosti.
 Druh byl objeven v roce 2000 na severu Madagaskaru při průzkumu v rezervaci Ankarana. Základní barva těla je skořicově hnědá. Srst na ocase je dlouhá a kolem očí vytváří tmavý prstenec. Stejně jako předešlý druh má tmavé vibrisy. Další z těch, kteří teprve čekají na důkladné prozkoumání. Díky výskytu v chráněné oblasti je výzkum poměrně slibný do budoucnosti.
Druh byl objeven v roce 2000 na severu Madagaskaru při průzkumu v rezervaci Ankarana. Základní barva těla je skořicově hnědá. Srst na ocase je dlouhá a kolem očí vytváří tmavý prstenec. Stejně jako předešlý druh má tmavé vibrisy. Další z těch, kteří teprve čekají na důkladné prozkoumání. Díky výskytu v chráněné oblasti je výzkum poměrně slibný do budoucnosti.
Microcebus simmonsi


Microcebus myoxinus


Microcebus rufus


Microcebus berthae


Microcebus lehilahytsara


Microcebus jollyae


Microcebus macarthurii
Vyskytuje se pouze v národním parku Makira na severovýchodě Madagaskaru. Známo velice málo informací.

Microcebus mittermeieri


Microcebus mamiratra


Microcebus bongolavensis
Je endemitem malé oblasti severního Madagaskaru a patří k větším druhům rodu. Známo málo informací.

Microcebus danfossi
Podobný předchozímu druhu. Vyskytuje se také na malém území o něco severněji.

Microcebus arnholdi
Rozeznán až v roce 2008 na základě genetických testů od druhu Microcebus sambiranensis. Je to maki tropického lesa s tmavou červenohnědou srstí na hřbetu a s kolísající váhou podle ročního období.
Microcebus margotmarshae
Popsán oficiálně také v roce 2008, vyskytuje se na severu Madagaskaru. Zbarven signifikantně oranžově s bílým nebo krémovým břichem.
Microcebus gerpi

Přináším originální popis tohoto druhu v angličtině.
Microcebus gerpi, sp. nova
Holotype. 002y09fina, adult male, captured on 25 June
2009, by B.R. and R.R. Tissue samples, hair samples as
well as e-voucher photos of the animal are stored at the
Institute of Zoology, University of Veterinary Medicine
Hanover, Germany. The animal itself was released after
field handling, sampling, and photographing, since its taxonomic distinctiveness was not recognized at the time of
capture. Field measurements (all lengths measured in mm):
ear length: 20.1, ear width: 11.2, head length: 35.3, head
width: 20.7, snout length: 7.6, intra-orbital distance: 7.4,
inter-orbital distance: 23.0, lower leg length: 40.1, hind
foot length: 22.8, third toe length: 8.9, body length: 85.0,
body mass: 77 g. The Sahafina population is designated the
source population for physical specimens in support of the
holotype.
Type locality: Forest of Sahafina (S18 47028.700,
E48 58056.200), in a savoka at about 95 m a.s.l. in the rural
community of Anivorano-East, Province of Toamasina,
Madagascar.
Paratypes: (a) 004y09fina, adult male, captured in the
Sahafina forest on 25 June 2009. Tissue and hair samples as
well as photos and morphometric measurements are stored
at the Institute of Zoology of the University of Veterinary
Medicine Hanover in Germany. (b) 003y09fina, adult male,
captured in the Sahafina forest on 24 June 2009. Tissue and
hair samples as well as photos and morphometric measurements are stored at the Institute of Zoology of the
University of Veterinary Medicine Hanover in Germany. It
is planned that one physical specimen will be obtained as a
further paratype in the near future, and that this specimen
will then be deposited in the Museum of the Zoology
Department of the University of Antananarivo, Madagascar.
Description: M. gerpi is a larger-bodied mouse lemur
with a brownish-gray body and a rufous-colored head. The
outer arms and legs are darker brown than the rest of the
body. The underfur is dense and short and covered with a
much sparser upper fur. The head is darker brown around
the eyes and has a distinct white stripe between the eyes
(Fig. 5). The ears are small but eminent and dark brown
towards the edges. The dorsum is grayish-brown with a
diffused and broad rufous line along the midline. The color
of the ventrum varies from a light gray to creamy white and
extends to the ventral throat and the genital region (including the scrotum). The tail is densely furred and long,
of brownish-gray color, and can store body fat (Fig. 5b). It
is darker on the dorsal than on the ventral side. Fingers and
toes are sparsely haired, and these hairs are whitish-gray.
The skin on hands and feet is pinkish-brown.
Diagnosis: M. gerpi can be distinguished from its closest
relative, M. jollyae, by genetic and morphological traits. It
has a significantly longer tail than M. jollyae and differs
genetically in 4.7% of its COII sequences and in 8.0–10.5%
of its D-loop sequences from its sister species. It can be
distinguished from its closest geographic neighbor, M. lehilahytsara, by its longer and broader head, its longer snout, its larger inter- and intraorbital distances, its longer
lower leg, third toe, tail, and body lengths, as well as its higher body mass. It differs genetically from M. lehilahytsara
in 6.0% of its COII sequences, in 11.2–12.3% of its D-loop sequences, and in 10.1–10.4% of its cytb sequences.
Furthermore, it differs from its northern neighbor, M. simmonsi, in 5.4% of its COII sequences, in 10.5–13.7%
of its D-loop sequences, and in 11.7–12.1% of its cytb
sequences. M. gerpi possesses ten diagnostic sites over all
three loci (4 bp in COII, 3 bp in cytB, 3 bp in D loop, and is therefore genetically distinct from all other mouse lemur species.
Etymology: M. gerpi is named after the Malagasy
organisation GERP (Groupe d’Etude et de Recherche sur
les Primates de Madagascar), which unites primatologists
working on Madagascar and is dedicated to research on and
the conservation of lemurs. All authors are members of GERP.
Vernacular name: English name: Gerp’s mouse lemur,
French name: Microce`be de Gerp, German name: Gerp’s
Mausmaki.
Notes: The distribution of M. gerpi is so far limited to
the Sahafina forest. This locality is not yet part of any
protected zone. Considering the presumably small distribution of this species and the present rates of deforestation in Madagascar, it is of the utmost importance to create a protected area for this species.
http://www.springerlink.com/content/v680g88l08tw6537/fulltext.pdf
 |
| a. paratyp, b. holotyp |
Rod Phaner
Phaner furcifer


Phaner pallescens
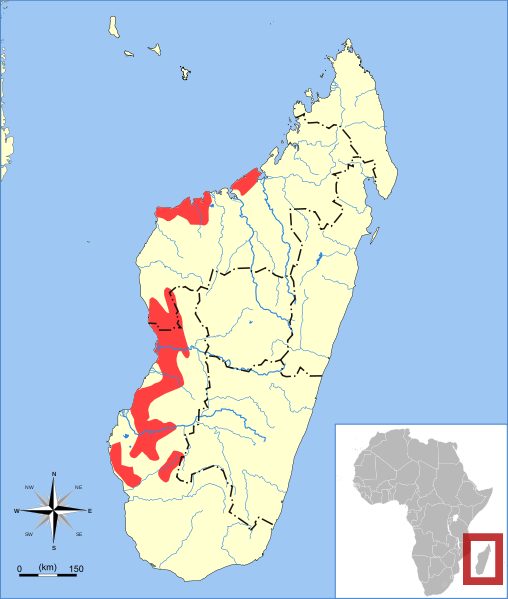
Phaner parienti
Málo známý druh ze severní části ostrova. V zbarvení podobný druhu Phaner furcifer, ale na úplném konci ocasu se kromě černé nachází také bílá špička. Není tolik vázán na mízu stromů.
Phaner electromontis
Zbarven spíše v sivém odstínu, najdeme ho na úplném severním cípu ostrova. Podobný druhu Phaner pallescens, ale ocas je tmavý více ke koncové části a černá linie hřbetu je více výrazná.

Rod Cheirogaleus
Cheirogaleus medius


Cheirogaleus major


Cheirogaleus crossleyi
Do rezava zbarvený maki s výskytem v několika vzdálených ostrůvcích původního lesa na východním Madagaskaru. Kolem očí ma typické tmavé kruhy. Jinak žije velice podobným životem jako předešlé druhy.

Cheirogaleus minusculus
Znám jen z lokality Ambositra ve středním Madagaskaru, jinak málo známý druh.
Cheirogaleus sibreei
Nezvěstný od roku svého popsání -1896. Znovu objeven v roce 2010, ale nikoliv v deštných lesích, kde se jeho výskyt předpokládal. Zdá se, že tenhle druh je velice ohrožen a vyskytuje se jen na omezeném území suchých lesů.
Jak je vidět, jediná čeleď lemurů se vyznačuje ohromnou diverzitou nejen tvarů a velikostí, ale také sociálních struktur nebo typů potravy. V dalších částech se zaměřím na jiné čeledi, u kterých mikroendemismus a adaptivní radiace také zanechali velice silnou stopu.
S radostí můžu oznámit, že v březnu 2012 byla publikována zajímavá studie o fylogenezi Cheirogaleidae, takže článek bude mít kvalitní dodatek a s aktuálními informacemi. V této studii je použit multilokusový data set, kde operační taxonomické jednotky (OTU) jsou nikoliv geny, ale jejich alely. Alignment byl tedy vytvořen na základě dat z jediné alely. Tenhle přístup ve skupině s mnoha kryptickými druhy a vysokou intraspecifickou diverzitou přinesl skvělé výsledky. Byla použita mitochondriální DNA a 12 genů jaderné DNA. Především zde najdeme první silně prokazatelnou pozici rodu Phaner, u kterého nebyli dosud výsledky jednotné. Tím je tedy vyřešena nejistá adaptivní radiace, o které jsem psal v úvodu. Phaner je sesterským kladem k ostatním cheirogaleidům, což bylo potvrzeno konsenzovými stromy a také Bayesiánskými stromy. U rodů Microcebus, Allocebus a Mirza byla nalezena diskordance mezi jednotlivými genovými stromy a autoři navrhují další studium vztahů mezi nimi. U Microcebus nedošlo k potvrzení nebo vyvrácení teorie o divergenci mezi suchými a deštnými lesy nebo té o rozdělení na severní a jižní skupinu. Naopak, najdeme zde výzvu k opatrnosti při interpretaci fylogeneze na základě analýzy stromu z jediného genu (s čím silně souhlasím). Celkově tato studie tedy ukazuje, že výběr alel může ovlivnit fylogenezi a čím více tedy použijeme lokusů, tím přesnější budou výsledky.
 |
| 4 z 10 stromů vytvořených na základě vyřezání jedné alely z jaderných genů. Červené hvězdy ukazují větve s podporou 95 %, což naznačuje malou konkordanci mezi těmito stromy. Faktor konkordance (CF) ukazuje počet genů, které dané větve podporují. |
 |
| Konsenzuální stromy pro 4 z 10 jaderných spojených data setů (majority-rule v Bayesu). Zde je vidět shodu v postavení rodů Phaner, Cheirogaleus, Allocebus a Mirza. V rámci Microcebus ale výběr různých alel způsobil diskordanci. |
Cheirogaleidae has probably the most complicated relationships between species among all lemur families. We include 5 genera here and two of them are a very mysterious ones for the phylogeny of this group. These small creatures with maximum weight a little more than 500 g are active during the night and this makes them a very difficult objects to study. For many species we just know that they exist. They are usually discovered on the very small area in case of their microendemism, but there is no time for collecting further data (it's time to change this). Moreover, we have no fossil evidence from this lemur family at all, what will be hopefully changed in the future (but it is very unlikely due to their fragile bones). The study of this interesting group is a field for new discoveries and we have to use only already known data about extand species.

The last very complex study of molecular evolution in Cheirogaleidae is from 2001 and it illustrates how much we still don't know about mouse lemurs. And in that time only 9 species were described, today we have 31 of them! Despite that we will take a look on this analysis. Already at that the two groups of karyotypes were discovered. In Phaner they found that 2n =46, in other four genera 2n=66. Phaner diversified probably as first from the common ancestor of this family and morphology supports this hypothesis (Tattersall and Schwartz, 1974). On the other side, imunological data from albumin and transferrin link Phaner and Cheirogaleus together and suggest Microcebus as the first to diversify. The analysis also revealed surprisingly early divergence of Cheirogaleus and sister group consisting of Microcebus and Mirza. Allocebus with only one species was not very well studied in that time so its position was unclear. Already in that time scientists suggested from mtDNA that new species of Microcebus will be discovered soon. In general we see, that Phaner and Cheirogaleus (possibly with Allocebus) were part of adaptive radiation and form a sister group to Microcebus and Mirza.
Cheirogaleidae probably diversified as the second group of lemurs after Daubentoniidae with only one species (Daubentonia madagascariensis). These tiny primates are often said to be a basal group very similar to first ancestors of whole order. From evolutionary point of view they really look primitive, but their social relationships are very complicated and we know soliters and also social species. A typical soliter is Mirza coquereli, Phaner furcifer and Cheirogaleus medius live in scattered pairs together with their offspring on particular area, in Microcebus murinus females form groups which spend the time when asleep together. A big role in formation of social structures was played by the enviroment (competition, predators, food availability).

Because the phylogenetic relationships between species of this family are not resolved, we will describe each of them individually. Analysis of mitochondrial DNA suggest the existence of two groups in genus Microcebus -eastern and western one. We can just wait for the discoveries of new species on new locations because of high level of diversification here.
Genus Allocebus
 The species has been considered to be extinct for a long time and was rediscovered as late as in 1989. It was assumed that it was critically endagered but its presence was proved on several areas of northeast Madagascar (rainforests). One of its typical characters is a long fur on its ears. The second and third upper molars are caniniform and the first upper incisor is enlarged. The shape of its ears is more similar to galagos than lemurs of Cheirogaleidae family. The tongue is rather long in comparison to other species of the family. It is a typical nocturnal species which sleeps in tree holes during the day. In dry season it increases its body weight with more fat reserves. Parents withstand their offspring in their territory. Its area is according to new research much bigger than in other tiny lemur species. The proper explanation for it haven’t been found yet although there was a suggestion that it is because of nutrition containing almost exclusively insect (some authors surprisingly claim that the main compund of the nutrition is tree gum) or seasonal changes of their behaviour. Allocebus doesn’t behave intolerantly even when sleeping in tree holes. From the evolutionary point of view it is at least an independent genus even though we couldn’t exclude the possibility of including it into an independent family (but there is no doubt that it is included deeply inside Cheirogaleidae according to molecular data).
The species has been considered to be extinct for a long time and was rediscovered as late as in 1989. It was assumed that it was critically endagered but its presence was proved on several areas of northeast Madagascar (rainforests). One of its typical characters is a long fur on its ears. The second and third upper molars are caniniform and the first upper incisor is enlarged. The shape of its ears is more similar to galagos than lemurs of Cheirogaleidae family. The tongue is rather long in comparison to other species of the family. It is a typical nocturnal species which sleeps in tree holes during the day. In dry season it increases its body weight with more fat reserves. Parents withstand their offspring in their territory. Its area is according to new research much bigger than in other tiny lemur species. The proper explanation for it haven’t been found yet although there was a suggestion that it is because of nutrition containing almost exclusively insect (some authors surprisingly claim that the main compund of the nutrition is tree gum) or seasonal changes of their behaviour. Allocebus doesn’t behave intolerantly even when sleeping in tree holes. From the evolutionary point of view it is at least an independent genus even though we couldn’t exclude the possibility of including it into an independent family (but there is no doubt that it is included deeply inside Cheirogaleidae according to molecular data). 
Genus Mirza

The Mirza family was originally included in Microcebus. It differs from it, however, with its size, weight up to 300 g and its way of life. It doesn’t hibernate in dry season but it stays active and feeds mainly on insect from order Homoptera. In other seasons of the year it also feeds on fruit and flowers. It is a typically solitary species. Sexual dimorphism was discovered in them, which is a great rarity among lemurs. Males are bigger and the size of their testicles is changing during the year (they mate with several females). The species appears only in dry forests of western part of the island ( the northern area belongs to a different species). Ears are big with little fur. Females have two pairs of nipples –pectoral and abdominal. Individuals of each sex build several nests in their home area which serve as shelters protecting them from predators. Lemurs of this species spend most of their time alone but a small part of it consists of social interactions such as mutual cleaning or vocalization.

Mirza zaza
This species was described only as late as in 2005. The description is based on behaviour studies and morphology. Genetic analyses confirmed its validity, the southern species was separated from the northern one approximately 2 million years. Mirza zaza is rather small, has smaller ears, shorter tail and canines. They differ in mitochondrial genom (cyt b) in 3,51%. Although the northern species is smaller, males have bigger testicles, which proves higher promiscuity. It is the biggest testicle size in comparison to the body size in all primates. The species differs also behaviorally – several individuals spend their time together in the nests ( there were even four adult males found in one nest).

Genus Microcebus

It is the first described (1777) and for a long time the only known species of Microcebus. Mouse lemurs belong to cryptic species living in a small area but this species is an exception – it appears in rather large area of western Madagascar in dry forest ecosystems. They are a typical nocturnal primates with big eyes and a long tail. We can find olfactory glands on their faces and around genitals. Females are bigger than males and the species is the biggest among genus Cheirogaleus with its weight up to 70 grams. In dry season they hibernate (torpor) with rapid decrease of vital functions and temperature only up to 7 °C while in other parts of the year they feed on different nutrition –from leaves and flowers to various insect. Females are dominant to males and also much more tolerant to other individuals of same sex. In the estrus females’ vaginas are not covered with fur as they usually are and males, otherwise solitary, form a hierarchy. The dominant male has, however, not guaranteed all the privileges because females mate with several males – we can therefore observe some kind of „arms race“ between the sexes. Whilst dominant males are mostly bulky and try to be superior to the others, the females tend to bring more genetic variability into the population, in other words, the „rare-male effect“ appears. These lemurs use a wide variety of acoustic (some of them
are even beyond human audability) and olfactoric signals for communication. The study of genetic variability of this species has revealed the existence of many others and a one-species genus became a genus of thirty species.

 Microcebus griseorufus
Microcebus griseorufusThis species lives in southern parts rather than the previous one and occupies the driest part of the island. It grows up to smaller size, it has light fur and more robust skull in comparison to the body. Study in the field suggested distinguishing of two species in this population based on morphological similarities. The mitochondrial DNA analysis, however, showed that there are no differences
between these two morphotypes. Such an unprecedented plasticity occurs in areas where mouse lemurs inhabit several microhabitats (a similar variability hasn’t been proved in areas with common occurence with the previous species). As nocturnal animals these primates don’t use visual communication as a primary source of information within social interaction, which explains the existence of two
different morphotypes. In the future, however, we may need to include nuclear genes in the analysis and might expect surprising results. But for now Microcebus griseorufus is a one species we still don’t know much about.

 |
| The two morphotypes from Beza Mahafaly locality |
Microcebus ravelobensis



A species from the northest part of the island where it inhabits the rainforest ecosystem. The coulour of the fur
consists of two or three different ones. The most significant one, however, is the dark strip stretching in the middle of its back. We can recognize dark whiskers on its head which are light among other species. It is one of the species we don’t know much about.


Microcebus sambiranensis
The species was discovered in 2000 in northern part of Madagascar during a research in Ankarana reservation. The basic colour of its fur is cinnamon brown. The fur on the tail is long and there are dark rings around its eyes. It has also dark whiskers as well as the previous species. It is another one from the group of species which are still waiting for a thorough research. Because of its location in a protected area the research is very promising.


It is the biggest one of genus Microcebus in the eastern part of Madagascar. It can weigh up to 75 grams. It appears only in Betampona reservation and Zahamena national park. The colouration is significant – from cinnamon to orange colour on its back with grey belly. There’s a distinctive white spot at the end of the rostrum. It was described as late as in 2006 during the study of mitochondrial DNA (D-loop) and morphometric data of Microcebus rufus species. Together with Microcebus simmonsi two other species were described.


Microcebus myoxinus
It is rather sorrel mouse lemur with its weight of approximately 30 grams, which makes it one of the world’s smallest primate. It lives in dry forests of north-western Madagascar. This species has much smaller ears, a longer tail and is a lot more agile than sympatrically living Microcebus murinus. During mating period the males are bigger than females but for the rest of the year it is contrary. This species don’t make any sleeping nests but they use of those produced by other lemur species, wholes in the trees or choose to sleep completely unprotected, which increases the risk of predation. Their social system is, like with other Microcebus species, based on male's promiscuity and sexual dimorphism.
It is rather sorrel mouse lemur with its weight of approximately 30 grams, which makes it one of the world’s smallest primate. It lives in dry forests of north-western Madagascar. This species has much smaller ears, a longer tail and is a lot more agile than sympatrically living Microcebus murinus. During mating period the males are bigger than females but for the rest of the year it is contrary. This species don’t make any sleeping nests but they use of those produced by other lemur species, wholes in the trees or choose to sleep completely unprotected, which increases the risk of predation. Their social system is, like with other Microcebus species, based on male's promiscuity and sexual dimorphism.


Microcebus rufus
It is a species widely spread on the whole of eastern Madagascar with more sorrel fur, shorter tail and smaller ears than Microcebus murinus. During the dry season we can observe their hibernation (torpor), which is, however, not always valid for males. They sleep in the tree holes during the day and occur mainly in ecotonal habitats, that means also in secondary forests. The species is principally fruitivorous but it enriches its food with insects. The male territories are bigger and often overlap several female territories. It is a solitary species which socializes with other individuals with a help of accustic and olfactoric signals (visual signals have also been described). This mouse lemur species belongs to the least endangered thanks to its wide occurance and an ability to survive in secondary forests. The peculiarity of this species is the length of its life – they live up to 8 years in the nature. This fact makes Microcebus rufus the shortest living primate in the world.
It is a species widely spread on the whole of eastern Madagascar with more sorrel fur, shorter tail and smaller ears than Microcebus murinus. During the dry season we can observe their hibernation (torpor), which is, however, not always valid for males. They sleep in the tree holes during the day and occur mainly in ecotonal habitats, that means also in secondary forests. The species is principally fruitivorous but it enriches its food with insects. The male territories are bigger and often overlap several female territories. It is a solitary species which socializes with other individuals with a help of accustic and olfactoric signals (visual signals have also been described). This mouse lemur species belongs to the least endangered thanks to its wide occurance and an ability to survive in secondary forests. The peculiarity of this species is the length of its life – they live up to 8 years in the nature. This fact makes Microcebus rufus the shortest living primate in the world.


It is the smallest primate of the world with a weight only of 30 grams. It occurs only in Kirindy reservation
in the west part of the island. Its fur is short and coloured in orange or yellow, being tawny overall. This mouse lemur originally belonged to Microcebus myoxinus but today it is considered to be a separate species living sympatrically with Microcebus myoxinus and Microcebus murinus. Male territories are larger than the female ones and as well as with other species of Microcebus genus, a promiscuous social system is assumed. A small area of this species is probably a result of a competition with Microcebus murinus, which has much fewer demands on the environment, while Microcebus berthae is bound to one single microhabitat. As regards food, Microcebus berthae is a lot more specialised than its bigger relative. Unlike its varied diet consisting of insect and its secretions, tree gum and fruit, Microcebus berthae feeds only on inscect secretions.


A species living only in Andasibe-Mantadia national park. It has a dark, reddish fur on the whole of its body and very dark circles around its eyes. There is a significant white spot on its nose tip. Microcebus lehilahytsara is a tree and nocturnal species, any other facts are, however, still unknown.


The species was described in 2006. It lives in two separate areas on the east of the island. Its colouring is sorrel, there is a significant white spot on its nose tip and and its belly is grey. Besides this there is only very little information known.

Microcebus macarthurii
It occurs only in Makira national park on the northeast of Madagascar. Not many facts are known
about this species yet.


It was also described in 2006 on the northeast of the island. This species is the smallest one of all from its genus in the eastern part of Madagascar with its weight of 40 grams only. The colour of its fur is mainly orange-brown. The species is seemingly very scarce because of extremely small area of occurence.

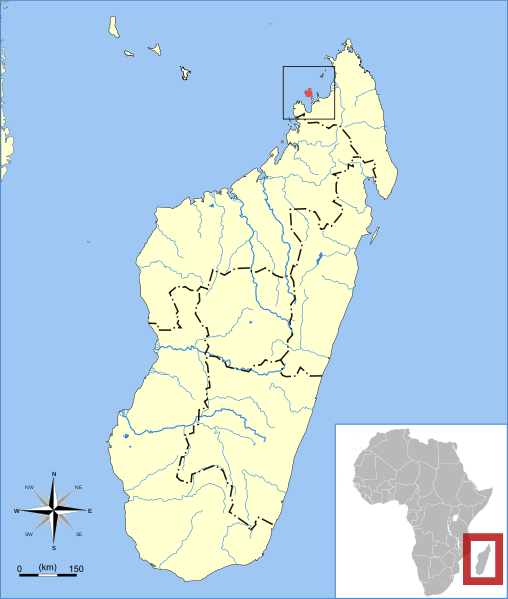 Microcebus mamiratra
Microcebus mamiratraIt lives on the island Nosy Bé on the northwest of Madagascar. Its colouring is sorrel and its closest relatives are Microcebus sambiranensis and Microcebus tavaratra. According to study of mitochondrial and nuclear DNA it differs from Microcebus sambiranensis only in the mitochondrial data, which may be a result of phylopatry of females (a return to their native area). Mitochondrial genom is inherited only from the mother and because the females stay in their native area even in their adulthood, the mitochondrial genomes of two separate populations differ. On the contrary, the nuclear DNA stays the same, therefore we can distinguish two subspecies, respectively two species in early stage of diversification.

It is an endemic species of a small area of northern Madagascar and it belongs to the bigger species of
this genus. Only little information is known about it.

Microcebus danfossi
It is similar to the previous species. It also occurs in the same area, only a bit more to the north.

Microcebus arnholdi
It was distinguished from Microcebus sambiranensis as late as in 2008 in pursuance of genetic tests. It is a mouse lemur of the tropical forests with dark sorrel fur on its back and a weight that fluctuates according to seasons.
Microcebus margotmarshae
Also officially described in 2008. It appears in the north of Madagascar. Its colouring is significantly orange with white or creamy belly.

The absolute novelty in the field of zoology! It was described only in 2012 after it was discovered that it varies from Microcebus lehilahytsara. Its area of occurence covers only a small territory on the east of Andasibe-Mantadia national park in the eastern part of Madagascar. This recently described species is significantly bigger with its weight up to 68 grams, as opposed to 46 grams of Microcebus lehilahytsara. Its closest relative is, according to genetic analysis of D-loop, cytb and COII, Microcebus jollyae appearing in territories on the south from Microcebus gerpi. Both of the species are sister taxons. Its body is rather massive and the tail can serve as a storage of fat. The fur of this species is brownish grey with red accents. Its ears are small in comparison to the body. This feature is typical for the species of tropical forests, as opposed to the ones living in dry forests on the north of the island (big ears in comparison to the body). There weren’t discovered any
differences between the sexes yet.
Here i provide the original description of this species.
Microcebus gerpi, sp. nova
Holotype. 002y09fina, adult male, captured on 25 June
2009, by B.R. and R.R. Tissue samples, hair samples as
well as e-voucher photos of the animal are stored at the
Institute of Zoology, University of Veterinary Medicine
Hanover, Germany. The animal itself was released after
field handling, sampling, and photographing, since its taxonomic distinctiveness was not recognized at the time of
capture. Field measurements (all lengths measured in mm):
ear length: 20.1, ear width: 11.2, head length: 35.3, head
width: 20.7, snout length: 7.6, intra-orbital distance: 7.4,
inter-orbital distance: 23.0, lower leg length: 40.1, hind
foot length: 22.8, third toe length: 8.9, body length: 85.0,
body mass: 77 g. The Sahafina population is designated the
source population for physical specimens in support of the
holotype.
Type locality: Forest of Sahafina (S18 47028.700,
E48 58056.200), in a savoka at about 95 m a.s.l. in the rural
community of Anivorano-East, Province of Toamasina,
Madagascar.
Paratypes: (a) 004y09fina, adult male, captured in the
Sahafina forest on 25 June 2009. Tissue and hair samples as
well as photos and morphometric measurements are stored
at the Institute of Zoology of the University of Veterinary
Medicine Hanover in Germany. (b) 003y09fina, adult male,
captured in the Sahafina forest on 24 June 2009. Tissue and
hair samples as well as photos and morphometric measurements are stored at the Institute of Zoology of the
University of Veterinary Medicine Hanover in Germany. It
is planned that one physical specimen will be obtained as a
further paratype in the near future, and that this specimen
will then be deposited in the Museum of the Zoology
Department of the University of Antananarivo, Madagascar.
Description: M. gerpi is a larger-bodied mouse lemur
with a brownish-gray body and a rufous-colored head. The
outer arms and legs are darker brown than the rest of the
body. The underfur is dense and short and covered with a
much sparser upper fur. The head is darker brown around
the eyes and has a distinct white stripe between the eyes
(Fig. 5). The ears are small but eminent and dark brown
towards the edges. The dorsum is grayish-brown with a
diffused and broad rufous line along the midline. The color
of the ventrum varies from a light gray to creamy white and
extends to the ventral throat and the genital region (including the scrotum). The tail is densely furred and long,
of brownish-gray color, and can store body fat (Fig. 5b). It
is darker on the dorsal than on the ventral side. Fingers and
toes are sparsely haired, and these hairs are whitish-gray.
The skin on hands and feet is pinkish-brown.
Diagnosis: M. gerpi can be distinguished from its closest
relative, M. jollyae, by genetic and morphological traits. It
has a significantly longer tail than M. jollyae and differs
genetically in 4.7% of its COII sequences and in 8.0–10.5%
of its D-loop sequences from its sister species. It can be
distinguished from its closest geographic neighbor, M. lehilahytsara, by its longer and broader head, its longer snout, its larger inter- and intraorbital distances, its longer
lower leg, third toe, tail, and body lengths, as well as its higher body mass. It differs genetically from M. lehilahytsara
in 6.0% of its COII sequences, in 11.2–12.3% of its D-loop sequences, and in 10.1–10.4% of its cytb sequences.
Furthermore, it differs from its northern neighbor, M. simmonsi, in 5.4% of its COII sequences, in 10.5–13.7%
of its D-loop sequences, and in 11.7–12.1% of its cytb
sequences. M. gerpi possesses ten diagnostic sites over all
three loci (4 bp in COII, 3 bp in cytB, 3 bp in D loop, and is therefore genetically distinct from all other mouse lemur species.
Etymology: M. gerpi is named after the Malagasy
organisation GERP (Groupe d’Etude et de Recherche sur
les Primates de Madagascar), which unites primatologists
working on Madagascar and is dedicated to research on and
the conservation of lemurs. All authors are members of GERP.
Vernacular name: English name: Gerp’s mouse lemur,
French name: Microce`be de Gerp, German name: Gerp’s
Mausmaki.
Notes: The distribution of M. gerpi is so far limited to
the Sahafina forest. This locality is not yet part of any
protected zone. Considering the presumably small distribution of this species and the present rates of deforestation in Madagascar, it is of the utmost importance to create a protected area for this species.
http://www.springerlink.com/content/v680g88l08tw6537/fulltext.pdf
 |
| a. paratype, b. holotype |
Genus Phaner

The genus Phaner grows up to a much larger size than the genus Microcebus. Phaner furcifer has a weight of approximately 300 g. It is a tree gum specialist –it has a tongue adjusted to gaining it and the bacteria in its gut help digesting this specific food. Its caniniform premolars are suitable to enrich the diet with insect. This typically colored species with reddish fur and a black stripe stretching along its back to its head occurs in the north-eastern part of Madagascar. It is typically nocturnal with territory area dependent on distribution of trees with enough tree gum. Phaner furcifer is a very dexterous climber in the treetops. The communication among individuals takes place mainly through acustic signals and unlike the Microcebus we can observe mostly monogamy here (polygyny is also possible).


A very similar species to the previous one with distribution in dry western part of Madagascar. It has less distinctive sorrel colour of fur and also the black stripe on its back. Phaner pallescens has a significant eyes lightning in the dark due a tapetum lucidum behind the retina.
Phaner parienti
Little known species from the northern part of the island. It has a similar coloration as Phaner furcifer but on the very end of its tail we can – besides the black colour- find also a white tip . It isn‘t bound to the tree gum so much.
Phaner electromontis
Colored in rather greyish tone, we can find it on the very northern tip of the island. It is similar to Phaner pallescens but the tail is dark near its end and the black stripe on its back is more significant.

Genus Cheirogaleus

A widely spread species in the dry west of the island. It is known for its hibernation when it withstands the dry season thanks to fat in its tail. Its coloriation is typically grey with black rings around the eyes, the ears are not coated. The hibernation lasts from 6 to 8 months. The young individuals start their first hibernation somewhat later, so as for them to collect enough
nutrients for the oncoming difficult period of time. One hole in a tree may be shared by several individuals. We can divide the active season into two separate ones – in the first one the lemurs feed mainly on nectar and fruit and in the second one they feed on insect or even small vertebrates. They are typically nocturnal lemurs with the peak of their activity after the sunset. They are, however, not as good climbers as species of genus Phaner and move in much slower manner. They communicate mainly in acustic but also in olfactory way. It is a monogamous species and the offspring stay in their parents’ territory. There are usually twins or triplets born.


It is bigger than the previous species, with reddish fur and a tail shorter than body. It hibernates and stores fat in its tail, too. This species occures widely and mainly in tropical forests of eastern Madagascar. It feeds mainly on fruit and nectar. Cheirogaleus major moves rather slowly and prudently. It is a solitary species and in the breeding season the individuals of opposite sexes communicate through vocalization.

Cheirogaleus crossleyi
A species with sorrel fur occuring in several distant islands with primary forests of eastern Madagascar. It has significant black rings around its eyes. Its lifestyle is similar to the previous two species.

Cheirogaleus minusculus
This species has only been observed in Ambositra locality in the middle of Madagascar. Otherwise very little known one.
Cheirogaleus sibreei
This species had been missing since 1986 - the year of its description. It was rediscovered in 2010, however, not in the rainforests where its distribution was assumed. This species seems to be highly endangered and lives only in a limited area of dry forests.
As we can see, a single family of lemurs can be characterized by a great diversity in morphology but also social structures or a type of diet. In the oncoming parts I will focus on other families where microendemism and adaptive radiation has left a very strong mark.
It is a pleasure to announce that in March of 2012 there was an interesting study of phylogeny of Cheirogaleidae published, therefore the article will have a apendix with current information. There is a multilocus dataset used in the study, where the operational taxonomic units (OTU) are not genes, but their alleles. The alignment was therefore based on data from one allele. This approach has brought a great results in a group with many cryptic species and high intraspecific diversity. A mitochondrial DNA and 12 genes of nuclear DNA were used. Above all, we can find the first strongly proved position of the genus Phaner, where the results hadn’t been integrated before. That solves the uncertain adaptive radiation which was mentioned in the introduction. Phaner is a sister clade to other cheirogaleids, which was confirmed by consenzus trees and Bayesian trees. In genera Microcebus, Allocebus and Mirza there was a discordance among individual gene trees found and the authors suggest further study of their relationships. The theory of divergence between dry forests and rainforests or the one of dividing it into the northern and southern group was neither confirmed nor refuted in genus Microcebus. On the contrary, we can find a call for caution with phylogeny interpretation based on the analysis of a tree from a single gene here (which I strongly agree with). On the whole, this study proves that the selection of alleles can influence the phylogeny and the more locuses we use the more accurate the results will be.
 |
| 4 of 10 trees created by cutting one allele of the nuclear genes. Red stars indicate branches supported by 95%, indicating little concordance between the trees. Concordance factor (CF) shows the number of genes that support the branch. |
 |
| Consensus trees for 4 of 10 nuclear concatenated data sets (majority-rule in Bayes). Here you can see the consenzus of position of Phaner, Cheirogaleus, Allocebus and Mirza. In Microcebus the selection of different alleles caused a discordance. |
{kind=link}
Zdroje (References) :
Jennifer Pastorini, Robert D. Martin, Petra Ehresmann, ElkeZimmermann and Michael R. J. Forstner. Molecular Phylogeny of the Lemur Family Cheirogaleidae (Primates) Based on Mitochondrial DNA Sequences. Molecular Phylogenetics and Evolution. Vol. 19, No. 1, April, pp. 45–56, 2001
doi:10.1006/mpev.2000.0904
Schülke, O. and Ostner, J. (2005), Big times for dwarfs: Social organization, sexual selection, and cooperation in the Cheirogaleidae. Evol. Anthropol., 14: 170–185. doi: 10.1002/evan.20081
Karla Biebouw. Home Range Size and Use in Allocebus trichotis in Analamazaotra Special Reserve, Central Eastern Madagascar. Int J Primatol (2009) 30:367–386
DOI 10.1007/s10764-009-9349-y
Bénédicte Viguier. Functional adaptations in the craniofacial morphology of Malagasy primates: shape variations associated with gummivory in the family Cheirogaleidae. Ann Anat 186 (2004}: 495-501
Christian Roos and Peter Kappeler. Distribution and Conservation Status of Two Newly Described Cheirogaleid Species, Mirza zaza and Microcebus lehilahytsara. Primate Conservation 2006 (21): 51– 53
Kappeler PM, Rasoloarison RM, Razafimanantsoa L, Walter L and Roos C. MORPHOLOGY, BEHAVIOUR AND MOLECULAR EVOLUTION OF GIANT MOUSE LEMURS (MIRZA SPP.) GRAY, 1870, WITH DESCRIPTION OF A NEW SPECIES. Primate Report 71, July 2005
Kellie L Heckman, Emilienne Rasoazanabary, Erica Machlin, Laurie R Godfrey and Anne D Yoder. Incongruence between genetic and morphological diversity in Microcebus griseorufus of Beza Mahafaly. BMC Evol Biol. 2006; 6: 98.
Published online 2006 November 16. doi: 10.1186/1471-2148-6-98
Sandra Thorén & Franziska Quietzsch &
Doreen Schwochow & Lalandy Sehen &
Christopher Meusel & Kate Meares & Ute Radespiel. Seasonal Changes in Feeding Ecology and Activity Patterns of Two Sympatric Mouse Lemur Species, the Gray Mouse Lemur (Microcebus murinus) and the Golden-brown Mouse Lemur (M. ravelobensis), in Northwestern Madagascar. Int J Primatol (2011) 32:566–586
DOI 10.1007/s10764-010-9488-1
Edward E. Louis Jr., Melissa S. Coles, Rambinintsoa Andriantompohavana, Julie A. Sommer, Shannon E. Engberg, John R. Zaonarivelo, Mireya I. Mayor and Rick A. Brenneman. Revision of the Mouse Lemurs (Microcebus)
of Eastern Madagascar. International Journal of Primatology, Vol. 27, No. 2, April 2006 ( C 2006)
DOI: 10.1007/s10764-006-9036-1
Dorothea Schwab and Jorg U. Ganzhorn. Distribution, Population Structure and Habitat Use of Microcebus berthae Compared to Those of Other Sympatric Cheirogalids. International Journal of Primatology, Vol. 25, No. 2, April 2004 (2004)
Melanie Dammhahn and Peter M. Kappeler. Comparative Feeding Ecology of Sympatric Microcebus berthae and M. murinus. INTERNATIONAL JOURNAL OF PRIMATOLOGY
Volume 29, Number 6, 1567-1589, DOI: 10.1007/s10764-008-9312-3
Ute Radespiel • Jonah H. Ratsimbazafy • Solofonirina Rasoloharijaona •
Herimalala Raveloson • Nicole Andriaholinirina • Romule Rakotondravony •
Rose M. Randrianarison • Blanchard Randrianambinina. First indications of a highland specialist among mouse lemurs (Microcebus spp.) and evidence for a new mouse lemur species from eastern Madagascar. Primates (2012) 53:157–170
DOI 10.1007/s10329-011-0290-2
www.theprimata.com
David W. Weisrock, Stacey D. Smith, Lauren M. Chan, Karla Biebouw, Peter M. Kappeler, and Anne D. Yoder. Concatenation and Concordance in the Reconstruction of Mouse Lemur Phylogeny: An Empirical Demonstration of the Effect of Allele Sampling in Phylogenetics. Mol Biol Evol (2012) doi: 10.1093/molbev/mss008
David W. Weisrock, Stacey D. Smith, Lauren M. Chan, Karla Biebouw, Peter M. Kappeler, and Anne D. Yoder. Concatenation and Concordance in the Reconstruction of Mouse Lemur Phylogeny: An Empirical Demonstration of the Effect of Allele Sampling in Phylogenetics. Mol Biol Evol (2012) doi: 10.1093/molbev/mss008
Ahoj,
ReplyDeleteměl bych pár komentářů; seřadím je od nejpodstatnějších k bezvýznamným: :-)
Poslední obsáhlá a komplexní studie ohledně molekulární stránky evoluce Cheirogaleidae je z roku 2001, což ukazuje, jak málo toho o trpasličích lemurech víme. Navíc v té době bylo popsáno jen 9 druhů, zatímco dnes jich známe 31!
Tohle mě zaujalo, takže jsem trochu hledal a našel jsem:
Weisrock DW, Smith SD, Chan LM, Biebouw K, Kappeler PM, Yoder AD 2012 Concatenation and concordance in the reconstruction of mouse lemur phylogeny: an empirical demonstration of the effect of allele sampling in phylogenetics. Mol Biol Evol doi:10.1093/molbev/mss008
Z abstraktu:
"In our study, we conduct a phylogenetic study for a nearly complete species sample of the dwarf and mouse lemur clade, Cheirogaleidae. Mouse lemurs (genus, Microcebus) have been intensively studied over the past decade for reasons relating to their high level of cryptic species diversity, and although there has been emerging consensus regarding the evolutionary diversity contained within the genus, there is no agreement as to the inter-specific relationships within the group. We attempt to resolve cheirogaleid phylogeny, focusing especially on the mouse lemurs, by employing a large multi-locus data set."
Zatím k tomu naneštěstí nemám přístup, ale kdyby ses k té práci náhodou nějak dostal sám, byl bych určitě moc vděčný za PDF -- ta studie má i hodně zajímavou metodologickou stránku (autoři konstruují species tree jako konsenzový strom z jednotlivých gene trees, jako OTUs používají jednotlivé alely).
Evolučně je to určitě alespoň samostatný rod, i když nelze vyloučit ani možnost zařazení do samostatné čeledi.
To je ale jenom věc vkusu; z dat je to nerozhodnutelné. V přírodě "rody" ani "čeledě" neexistují, vymysleli jsme si je my čistě pro usnadnění komunikace, a ani tenhle účel nijak zvlášť dobře neplní. Poslední pokus objektivně definovat ranky, aby šlo o těchhle otázkách nějak smysluplně rozhodovat, tady byl za Sibleyho a Ahlquista -- přičemž tihle chudáci ještě věřili ve striktní molekulární hodiny (ve stylu "1,5% rozdílné DNA ~ divergence před 5 miliony let") a neznali problémy s alignmentem.
[...] a také oblasti, kam se upínají (koronoidní processus).
Proč ne prostě "koronoidní výběžek"? Docela běžně se to používá.
Cheirogaleidae se zřejmě odštěpili od hlavní fylogenetické linie lemurů jako druzí hned po Daubentonidae s jediným zástupcem, ksukolem ocasatým.
Chápu, jak to myslíš, a vím, že není úplně jednoduché tuhle formulaci obejít, ale fylogeneze samozřejmě žádné hlavní linie nemá. Ani u lemurů, ani nikde jinde. :-)
Ahoj,
ReplyDelete1. taky mně to docela sklamalo a tohle je od tebe skvělej objev, za který jsem ti vděčnej:) Samozřejmě to vyšlo těsně před tím než jsem psal článek a zřejmě to ještě nebylo zahrnuto v databázích, proto mi to uniklo. Takže teď se dočkáme dodatku ke článku.
tady to je, doufám že ti to půjde otevřít:)
http://mbe.oxfordjournals.org/content/early/2012/03/23/molbev.mss008.full.pdf+html
2.Samozřejmě je to jen věc vkusu, což vidíme všude kolem a právě kvůli tomu je systém s čeleděmi, rody a tak dále občas nepřehlednej a nedá se srovnávat čeleď a čeleď nebo rod a rod. Já to používám právě jen tehdy, když to může čtenáři napomoct se zorientovat. Kdybych psal jenom a výhradně o kladech, tak je nutno mít opravdu přehled o situaci v dané skupine živočichů.
3. Nevím, prostě mi to přišlo moc jednoduché:D
4.Přesně tak, těžko se to obchází a když si vezmeš, že se ksukol oddělil od všech ostatních lemurů, tak hlavní linie se nabízí jako možné pojmenování pro vysvětlení(samozřejmě bez dalšího významu).
Znovu ahoj,
ReplyDeleteomlouvám se za dlouhou prodlevu.
tady to je, doufám že ti to půjde otevřít:)
http://mbe.oxfordjournals.org/content/early/2012/03/23/molbev.mss008.full.pdf+html
Naneštěstí nejde, mám to za paywallem. Myslíš, že bys mi mohl poslat kopii na email? Byl bych ti vážně vděčný... ;-)
Já to používám právě jen tehdy, když to může čtenáři napomoct se zorientovat.
Ehm... snad do toho nebudu přehnaně šťourat, ale co přesně si má čtenář odnést z toho, že "taxon X je minimálně samostatný rod, možná i čeleď", když sami přiznáváme, že neexistuje objektivní způsob, jak to rozhodnout? Napadly mě tři scénáře:
1) Nevíme, zda je Allocebus maki, tzn. jestli je blíž příbuzný cheirogaleovi než ostatním lemurům. Byly tady nějaké pochybnosti tohohle typu? Z tvých fotek mi připadá, že mezi ty ostatní chlupáče zapadá celkem dobře...
2) Allocebus by mohl být sesterská skupina všech ostatních makiovitých. Jenže pak je fakt jen na nás, jestli mu tu vlastní "čeleď" dáme nebo ne; Cheirogaleidae budou monofyletičtí tak jako tak, jde jen o to, jak široký bude jejich rozsah. Je to prostě klasická otázka splitting vs. lumping; občas se ujme to první (třeba odštěpení pozemních zoborožců z Bucerotidae do Bucorvidae), jindy to druhé (pokus vztyčit Dendrocygnidae a omezit rozsah Anatidae nevyšel, i když s fylogenezí, na které byl založen, většina lidí souhlasí). Je to vážně jen věc vkusu a konsenzu; jak to může pomoct s orientací, nevím.
(Omlouvám se za příklady ilustrované na ptácích, ale v placentálech se nevyznám.)
3) Skutečně používáš taxonomickou úroveň jako měřítko distinktivnosti, tj. na fylogenetické pozici zas tak moc nezáleží, důležité je, že Allocebus je oproti ostatním makiům prostě divný. Dobře, ale vážně to chceme zdůraznit v taxonomii? Stojí to za ty problémy? Podle těch fylogramů výše to vypadá, že Allocebus je v rámci makiovitých docela hluboce usazený, a nezapomínejme, že ICZN neumožňuje mít jednu "čeleď" zanořenou do druhé. Takže se buď smíříme s tím, že Cheirogaleidae budou parafyletičtí, nebo je roztrháme na menší, monofyletické "čeledě". V obou případech si úplně zbytečně komplikujeme život, v tom druhém případě by to navíc znamenalo umístit 5 "rodů" do 4 "čeledí", což taxonomii úplně absurdně nafukuje.
Pointa: nejenže nevím, co z toho by "nelze vyloučit zařazení do samostatné čeledi" mohlo znamenat, ale vždy existuje alternativa, jak to říct líp: "není jisté, jestli je to vůbec maki"; "může být sesterským taxonem vůči všem ostatním makiům"; "je to hodně divný maki".
Ahoj,
ReplyDeletestudii máš na emailu:)
Zcela s tebou souhlasím, že to šlo říci i jinak:) Allocebus měl kdysi zcela nejasné postavení a spekulovalo se o tom, kam ho zařadit (rozdílná dentice a další morfo znaky). Až molekulára postupně ukázala, že sedí někde ve středu Cheirogaleidae, i když na základě třeba etologických a behaviorálních odlišností by se našli takoví, kteří by mu vytvořili samostatnou čeleď. To bylo ale dříve, když to vypadalo tak, jak píšu na začátku (polytomie, kam patřil i Allocebus). A právě z důvodů, které píšeš, je prostě mezi Cheirogaleidae v samostatném rodu. Stejně jako u Dendrocygna by se to prostě neujalo (nevadí mi jakýkoliv příklad z obratlovců) :).
Abych byl stručný, Allocebus je poněkud zvláštní maki, ale dnes už ho vidíme díky molekulárním datům v Cheirogaleidae. :)
Jinak je zajímavé sledovat, jak rozdílně se splitting uplatňuje u jiných skupin. Především u ptáků je království splitterů, kde se systém pořád mění a překopává (budu psát třeba o Mesitornithiformes). U savců je tomu zase naopak, kdyby byl třeba Allocebus zařazen do samostatné čeledi, asi by z toho byl docela rozruch. Jak to vidíš ty, co se týče těch rychlých změn u ptáků?
Ahoj, studii máš na emailu:)
DeleteJeště jednou díky.
Především u ptáků je království splitterů, kde se systém pořád mění a překopává
Docela zajímavé je, že situace je dost odlišná u pěvců a non-pěvců -- jak hezky ukázal třeba Harshman (2007). U non-pěvců jsme si nejsme jistí monofylií "řádů", ale "čeledě" jsou monofyletické docela určitě (až na pár výjimek v trubkonosých a dlouhokřídlých) a jednotlivé taxony nám nepřeskakují z jedné do druhé. Zato pěvčí "čeledě" jsou hodně nestabilní a "rody" mezi nimi přeskakují docela běžně. U non-pěvců se vlastně názvosloví poslední 4 roky jen vyrovnává s tím, s čím přišla fylogenomika: fregatky, terejové, kormoráni a anhingy už nejsou Pelecaniformes, ale Suliformes; jestřábovití, orlovec a hadilov (a možná i kondoři) už nejsou Falconiformes, ale Accipitriformes. U pěvců je toho daleko, daleko víc: z mravenčíkovitých se odštěpili Grallariidae, Sylviidae se rozpadá už dlouho, naposledy je opustili tuším že budníčci (Phylloscopidae), Dendrocoleptidae zase "čeleď" ztratili ve prospěch Furnariidae (kteří by vůči nim jinak byli parafyletičtí), pochyby jsou ohledně dalšího statusu Acanthizidae atd. ad nauseam.
budu psát třeba o Mesitornithiformes
Jo, tak tohle třeba vůbec nechápu. Co je špatného na "Mesitornithidae"? Touha, aby každý pták patřil pod právě jedno jméno s koncovkou "-iformes", musí být nějaká divná úchylka systematické ornitologie.
BTW, když jsme u divných madagaskarských ptáků, kurol by si taky zasloužil svých pět minut slávy! Navíc by to šlo skrze pleziokatarta výborně propojit se sérií o Messelu.
Jak to vidíš ty, co se týče těch rychlých změn u ptáků?
Řekl bych, že ornitologům je ve většině případů prostě jen líto odvrhovat stará jména kvůli novým fylogenezím. V mnoha případech by bylo nejvhodnější se některých "čeledí" prostě zbavit (lumping): třeba když se ukázalo, že vousáci jsou masivně parafyletičtí vůči tukanům (což není nijak nové -- ví se to minimálně od 80. let a ani k tomu nebyla potřeba molekulára) nebo že tenkozobci, ústřičníci a srpatky jsou blíž příbuzní kulíkům a čejkám, než někteří další "kulíci" (Pluvianellus, Pluvialis). V takových situacích by bylo nejlepší ponořit Ramphastidae do Capitonidae a Recurvirostridae, Haematopodidae a Ibidorhynchidae do Charadriidae. (Mimochodem, "Haematopodidae" a "Ibidorhynchidae" jsou stejně úplně zbytečnými synonymy příslušných "rodů".) Ornitologové to ale neudělali a radši se vydali cestou rozdrobování parafyletických "čeledí" do menších monofylů, takže vousáci už byli rozdrobeni na Megalaimidae, Lybiidae, monotypické "Semnornithidae" a nakonec staré dobré Capitonidae, tentokrát ale omezené na americkou podskupinu -- to vše jen proto, abychom mohli zachovat Ramphastidae. S kulíky to jde daleko snáz (tj. prostě přestat zahrnovat pluvialise do Charadriidae), ale některým to nestačilo: všichni ptáci by nejspíš okamžitě vymřeli, kdyby nějaký z nich nepatřil do žádné "čeledě", takže je potřeba nějakou rychle zavést pro každý jednotlivý rod, který v nových fylogenezích svou tradiční "čeleď" opustil. Odtud třeba absurdní systematika Bakera a spol. (2007), rozeznávající vedle sebe "Pluvialidae", "Pluvianidae" a "Pluvianellidae". Přeji snadnou orientaci v kladogramu.
Odkazy:
DeleteBaker AJ, Pereira SL, Paton TA 2007 Phylogenetic relationships and divergence times of Charadriiformes genera: multigene evidence for the Cretaceous origin of at least 14 clades of shorebirds. Biol Lett 3(2): 205–9
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2375939/pdf/rsbl20060606.pdf
Hackett SJ, Kimball RT, Reddy S, Bowie RC, Braun EL, Braun MJ, Chojnowski JL, Cox WA, Han K, Harshman J, Huddleston CJ, Marks BD, Miglia KJ, Moore WS, Sheldon FH, Steadman DW, Witt CC, Yuri T 2008 A phylogenomic study of birds reveals their evolutionary history. Science 320(5884): 1763–8
http://www.luc.edu/biology/reddy/hackett2008.pdf
Harshman J 2007 Classification and phylogeny of birds. 1–35 in Jamieson BGM, ed. Reproductive Biology and Phylogeny of Birds. Enfield (NH): Science Publishers Inc.